


Most of the plants we see around us today arrange their leaves, flowers or seeds in a special pattern that forms spirals based on the Golden Angle (137.5°) associated with the Fibonacci mathematical series. This precise arrangement is thought to optimize both space and sunlight for the plant’s organs. It is also ancient – making an appearance in the fossil record about 380 million years ago during the Late Devonian, if not earlier.
Striking examples of modern Fibonacci spirals include sunflower heads, succulents' stems and leaves as well as conifer seed cones.



Left to right: Spiral phyllotaxis in sunflower (Helianthus annuus) by explorer2023 (CC BY-NC 4.0); Spiral Aloe (Aloe polyphylla) by juddkirkel (CC BY-NC 4.0); and conifer cones by harum.koh (CC BY-SA 4.0). Images from iNaturalist.
“Fibonacci spirals constitute 91 % of documented phyllotactic patterns in 650 species of angiosperms and gymnosperms, although this type of phyllotaxis is also found in ferns, clubmosses, spike mosses, quillworts and mosses,” write Howell and colleagues in their new paper recently published in a special issue of Annals of Botany, “Role of Fossils in Reconstructing Plant Evolution”.
Historically, scientists have thought that because this trait is so prevalent amongst plants, it must be the ancestral state for plant phyllotaxis. But recent evidence from Asteroxylon mackiei, a lycopod of the Early Devonian, complicates the picture, having “both whorls and non-Fibonacci spirals”, indicating that Fibonacci spirals might have evolved later in leafy plants.
This finding then opens the question of how form affected function as spiral phyllotaxis evolved.

And so, Howell and colleagues set out to analyse the spiral phyllotaxis of seed cones of the Middle Jurassic using 165 million year old seed cones of the conifer Araucaria mirabilis and test whether the conifer cone attributes affected seed packing.
The researchers used non-destructive micro-CT scanning as well as gross morphological measurements to ask whether cone axis, cone proportions (length, width, circumference), chirality (handedness) or parastichy attributes affected the number seeds in the cone (seediness).
Twenty-one well-preserved A. mirabilis seed cones, originally from the Bosques Petrificados de Jaramillo National Park in Patagonia, Argentina, were selected for cone analysis from the paleobotanical collection at the Museum für Naturkunde Chemnitz in Chemnitz, Germany.
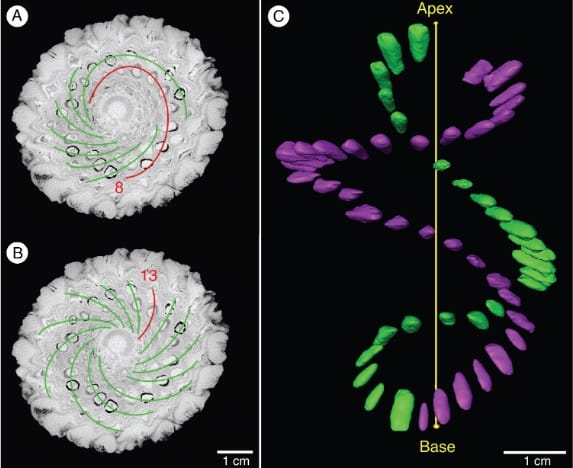
Overall, they found that having a longer cone axis and a clockwise spiral is associated with increased cone seediness, potentially being key to optimal cone packing.
“Based on the correlations observed in this study, clockwise ontogenetic chirality is an advantageous feature of A. mirabilis seed cones,” write Howell and colleagues. The clockwise orientation was correlated with tighter spirals and more seeds.
By performing this detailed study on cone spiral phyllotaxis, Howell and colleagues were able to document the biological functions of these patterns for the first time and show how they relate to seed packing in ancient – and perhaps modern – conifers.
READ THE ARTICLE:
Howell, M., Rößler, R., and Gee, C.(2025) Phyllotaxis, ontogeny and CT imaging: old and new approaches to understanding optimal seed packing in Middle Jurassic Araucaria mirabilis cones. Annals of Botany, 137(6), pp. 1817-1831. Available at: https://doi.org/10.1093/aob/mcaf325.


