


Many questions about plant development can be answered through direct observation. Others, such as those involving the effect of molecular-level interactions on development and form, also require computational modelling.
Dr. David Holloway and Dr. Carol Wenzel, both at the British Columbia Institute of Technology, use plant modelling to investigate the formation of the multi-vein secondary network in dicot leaves in their recent in silico Plants article.
The authors developed a leaf-level model that simulates intracellular and membrane kinetics and intercellular transport. With it, they were able to analyze the dependence of vein patterning on the efficacy of auxin flux through cells.
The phytohormone auxin is involved in regulating plant growth and drives vascular development, forming veins as it flows through the leaf. The direction of auxin flow is regulated by PIN-FORMED cell efflux transporters. These transporters are localized to specific sides of cells and promote the pumping of auxin molecules outside cells, directing flow. There appear to be two general trends in transporter-auxin interaction: an up-the-gradient transporter allocation where auxin moves toward neighboring-cells with high auxin concentrations, and a with-the-flux allocation dependent on the flow of auxin across walls and away from high auxin concentrations.
The authors combined up-the-gradient and with-the-flux allocation to create a dual-polarization auxin transport model. This mechanism successfully simulates normal primary and secondary vein development. This includes transporter-auxin distributions corresponding to single-vein initiation, extension and canalization, as well as the synchronized and sequential initiation of secondary veins, control of their extension direction, and connection of the secondaries to the mid-vein (see figure 1).
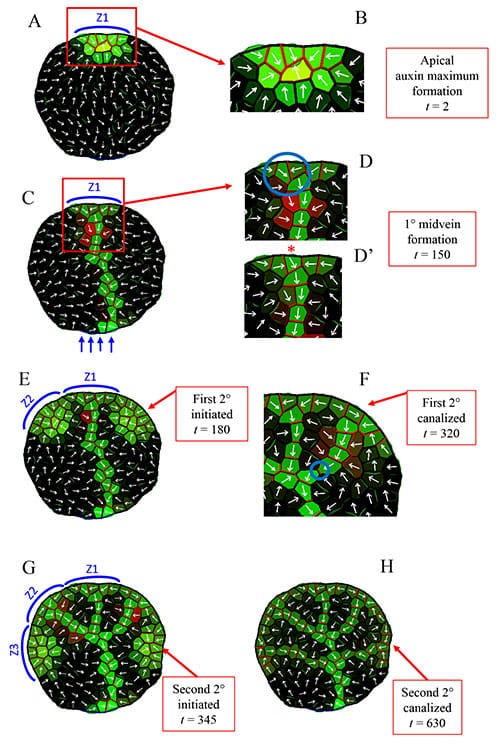
“After the static template was found to be successful, we integrated cellular growth and division. From this, we were able to show how growth can expand early venation patterns from the cellular resolution (where molecular dynamics play a role) to the whole leaf” says Holloway.
The role of auxin transporters in vein extension was further investigated by modelling the chemical inhibition of auxin transport. Simulated moderate and high levels of inhibition corresponded well with what is observed experimentally:
Moderate inhibition resulted in a switch from several distinct vein initiation sites to many less-distinct sites; a delay in vein canalization; inhibited connection of new veins to old; and finally loss of patterning in the margin, loss of vein extension, and confinement of auxin to the margin (see figure 2A vs 2B). High inhibition resulted in more convergence points of high auxin levels forming in the margin than normal, but without initiating extending veins (see figure 2A vs 2C).
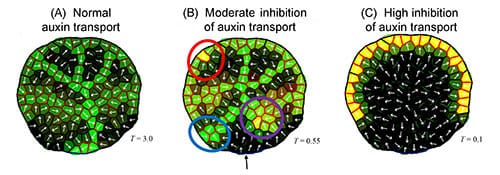
Holloway concludes “modelling normal and transport-inhibited leaves allowed us to identify dynamics involved in auxin directional flow changes during development of the secondary vein network. We found that cellular-scale auxin transport produces short-range vascular patterns, and demonstrated that leaf growth can be a simple mechanism for expanding short-range patterns to the scale of the mature leaf.”
READ THE ARTICLE:
David M Holloway, Carol L Wenzel, Polar auxin transport dynamics of primary and secondary vein patterning in dicot leaves, in silico Plants, 2021; diab030, https://doi.org/10.1093/insilicoplants/diab030




