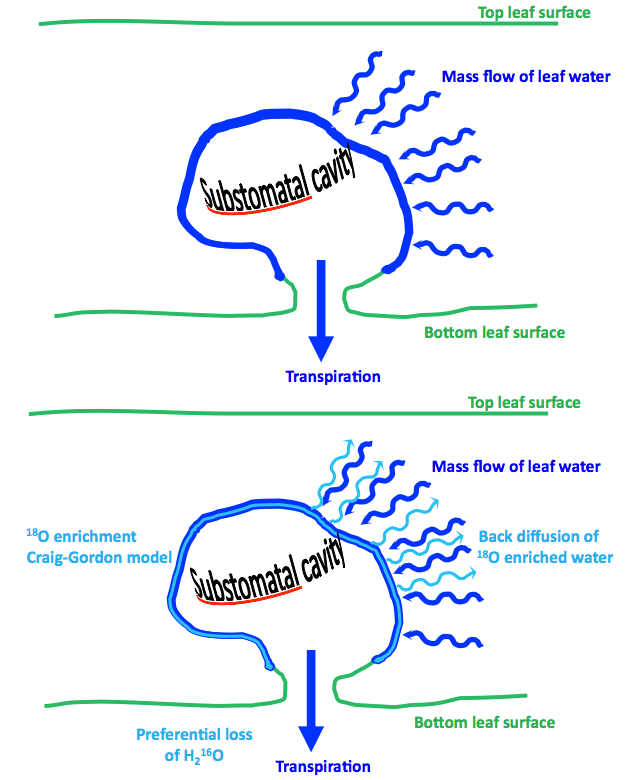


Stable oxygen isotope analyses have various applications relevant to tracking water movement in ecosystems. The ratio of 16O to 18O (represented as δ18O) in leaf water provides information about the water vapor pressure deficit (VPD) in the air around the plant, the plant’s source of water and the physiological processes that govern leaf water loss, such as stomatal movements and transpiration. For example, as VPD increases (i.e. as environmental conditions become drier), more evaporation and thus evaporative enrichment in 18O occurs within the leaf water, since the lighter H216O evaporates more readily than the heavier H218O. This causes the remaining H2O at the sites of evaporation to become more enriched in 18O. Researchers have used this model, originally applied to oceans by Craig and Gordon (1965), to shed light on evaporative conditions and leaf physiology.
However, the Craig-Gordon model resulted in overestimates of leaf water H218O enrichment because it did not account for the less enriched water flowing from leaf veins to the sites of evaporation. This overestimation greatly impacts how we interpret δ18O as a proxy for evaporative conditions in which the plant grew, as well as leaf physiology. In response to this overestimation, Farquhar and Lloyd (1993) proposed the Péclet effect defined as: water flowing towards sites of evaporation by transpiration becomes enriched by the back diffusion of the enriched water at the sites of evaporation. In other words, the Péclet effect describes the mixing of water from both the xylem and evaporative sites. The Péclet effect is largely driven by changes in transpiration rate (E) and effective path length (L), where L describes the tortuous path that water travels from leaf veins to sites of evaporation.

To better inform interpretations of leaf water δ18O, the Craig Gordon-Péclet model is used to predict L. L is impossible to measure because it accounts for not only the distance that water travels, but also the tortuosity of the water movement pathway. However, the model requires assumptions about evaporative conditions that may or may not be true.
Previous work (Song et al. 2013) documented relationships between L and physiological parameters such as E and hydraulic conductance (k). However, few studies have estimated L under controlled environmental conditions. Loucos and colleagues did just that, and estimated L and k simultaneously on the same leaf to evaluate the findings of previous studies.
Loucos et al. (2015) found that estimates of L are strongly influenced by assumptions made when calculating 18O enrichment at the sites of evaporation. Contrary to previous studies, they found no support for the hypothesis that L is negatively related to both E and k within a single species. This negative correlation was expected because water movement was expected to follow a predominately small L pathway when E is high (i.e. E > 1 to 2 mmol m-2s-1), and when E is low (i.e. E < 1 to 2 mmol m-2s-1), there is a proportionally greater flux through a large L pathway (Song et al. 2013). As a result, the authors demonstrate that great caution must be taken when investigating and developing any relationships between L and environmental and physiological parameters. Clearly, the path to understanding L, the Péclet effect, and δ18O of leaf water is not straightforward.
References
Craig H. & Gordon L.I. (1965). Deuterium and oxygen-18 variations in the ocean and the marine atmosphere. In: Tongiorgi E (ed)
Proceedings of a conference on stable isotopes in oceanographic studies and paleotemperatures,.Laboratory of Geology and Nuclear Science, Pisa, pp 9-130.
Farquhar G.D. & Lloyd J. (1993). Carbon and Oxygen Isotope Effects in the Exchange of Carbon Dioxide between Terrestrial Plants and the Atmosphere. In: Ehleringer J.R., Hall A.E., Farquhar G.D. (eds)
Stable Isotopes and Plant Carbon-water Relations.Academic Press, San Diego. 47-70. DOI: http://dx.doi.org/10.1016/b978-0-08-091801-3.50011-8
Loucos K.E., Simonin K.A., Song X. & Barbour M.M. (2015). Observed relationships between leaf H218O Peclet effective length and leaf hydraulic conductance reflect assumptions in Craig-Gordon model calculations,
Tree Physiology,DOI: http://dx.doi.org/10.1093/treephys/tpu110
Song X., Barbour M.M., Farquhar G.D., Vann D.R. & Helliker B.R. (2013). Transpiration rate relates to within- and across-species variations in effective path length in a leaf water model of oxygen isotope enrichment,
Plant, Cell and Environment, 36(7) 1338-1351. DOI: http://dx.doi.org/10.1111/pce.12063



